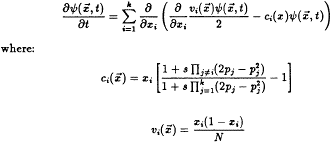![]()
|
Punctuated Equilibrium Due to Epistasis
in |
|||
|
|||
|
Abstract
|
|||
|
experimental parameter. At this point, randomized
point mutations or transpositions may
|
|
eliminated. A third method of selection is to
assign an a priori probability of survival to each |
||
|
|
||
|
|
||
|
|
|
selective advantage. For a finite randomly mating
population of size N, the evolution of the |
||||
|
|
||||
|
|
||||
|
|
||||
|
For small values of s, this may be closely approximated by: |
||||
|
|
||||
|
|
||||
|
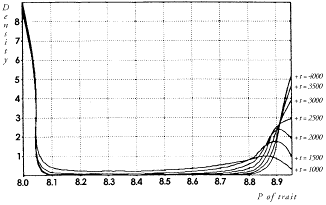
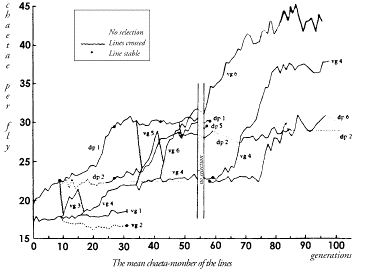
small increase in the frequency of one allele
will greatly enhance the selective values of the |
|||
|
|
|||
|
|
|||
|
|
|||
|
|
|||
|
|
|||
|
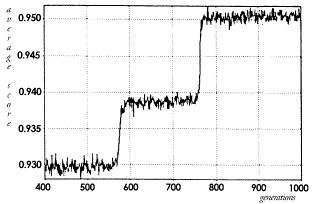
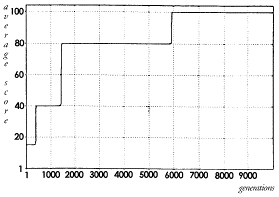
Discussion
|
||
|
|
||
|
|
||
|
Acknowledgements
|
|
Tejeda, A.L. "Impact of the Use of Mixtures
and Sequences of Insecticides in the Evolution
|
To bookmark this page click here and then bookmark this unframed version.